Engineering
Our project Electricia coli is based on the use of a genetically modified Escherichia coli as a biosensor
that
emits an electrical signal when detecting a specific cancer biomarker: the PANTR1 long non-coding RNA (lncRNA).
Our Electricia coli biosensor is based on the use of a toehold switch [1] (Figure 1). This RNA-based device
is
composed of several parts which are essential for the efficient functioning of our biosensor:
- a Trigger Binding Site (TBS)which is a sequence complementary to the RNA target to be detected
- a Ribosome Binding Site (RBS)which allows the start of the translation only when the toehold switch is unfolded
- a start codon AUGwhich constitutes the start of the translation
- a linkerwhich constitutes a bridge between the Start codon and our gene of interest
- a gene of interestthat is translated into a protein only when the hairpin is unfolded and the RBS exposed
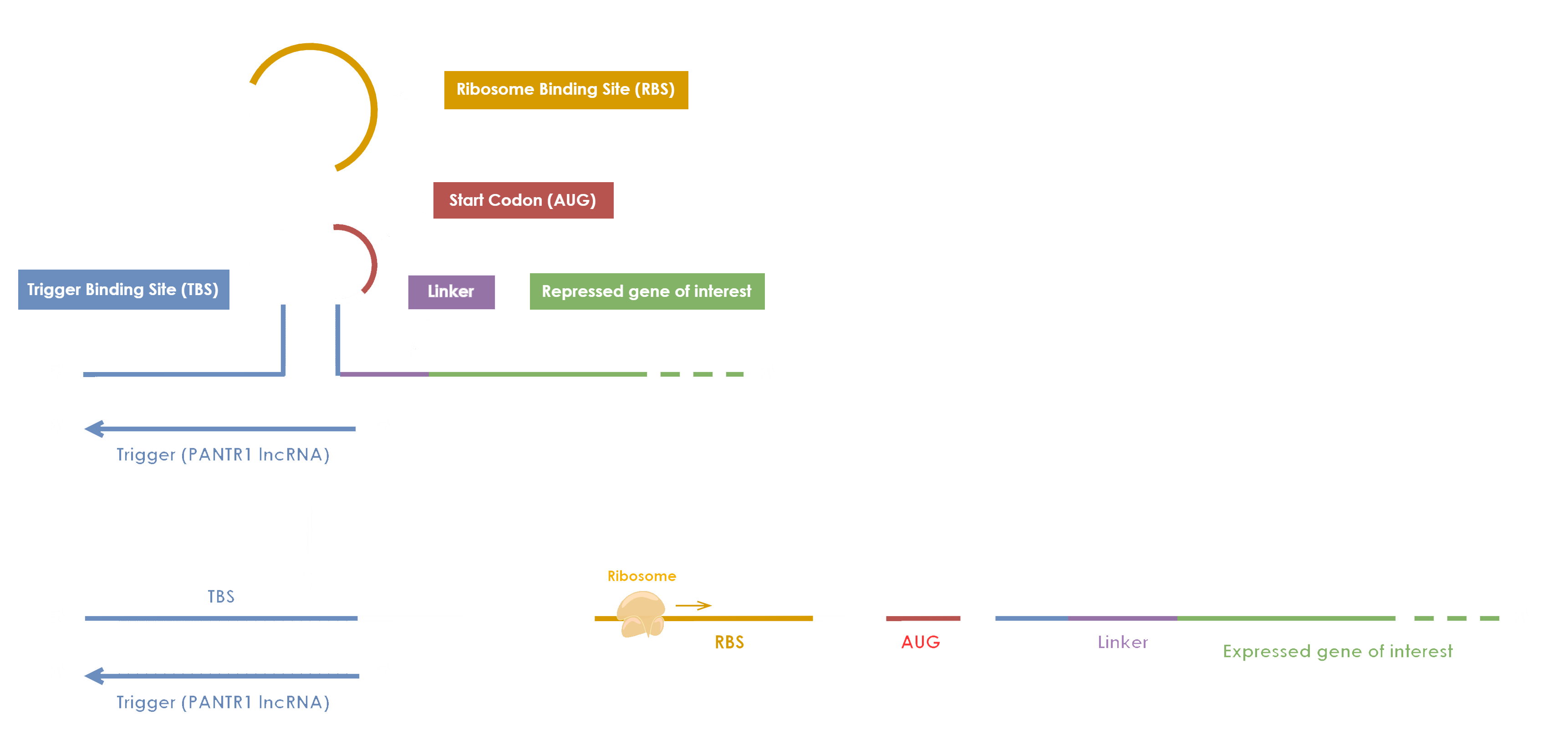
Figure 1. Principle of the toehold switch used for Electricia coli (inspired from iGEM Evry
Paris-Saclay 2020).
The functioning comes as follows:
The binding between the RNA biomarker and the toehold switch unfolds the hairpin structure. The RBS becomes accessible now so that the translation can take place, and the initially repressed gene of interest can then be translated into a protein. In the Electricia coli project, the translated protein plays a role in the production of electrons. By using an MCF, electrons are captured and converted into an electrical signal, which is recorded by an embedded system.
We were inspired by previous such biosensors designed to specifically identify various biological nucleic acid sequences originating from viruses [2–7], trees, [8] or cancer related mutations [9], and applied the DBTL (Design, Build, Test, Learn) framework, a hallmark of synthetic biology, in order to develop high performing toehold switches as biosensors that detect the PANTR1 lncRNA.
Electricia coli “Design – Build – Test – Learn” Cycle
Design
Our aim was to give E. coli the ability to detect the lncRNA PANTR1. To do so, we have to design some toehold switches with computational approaches, and test in vivo the ones with the best performance. We use superfolder green fluorescent protein (sfGFP) as the gene of interest for easy screening of protein expression.The prediction of the toehold switches was made with the tool developed by the Hong Kong-CUHK iGEM 2017 team [10] for each of the four versions of the PANTR1 lncRNA. We choose to adopt the previously optimized architecture of the toehold switch sensors for Zika virus detection [2] that proved its efficiency on other toehold switches too [8,11]. This architecture comprises a custom Loop sequence (GGACUUUAGAACAGAGGAGAUAAAGAUG) and a custom Linker sequence (AACCUGGCGGCAGCGCAAAAG), which we specified as parameters in the CUHK web tool [10].
A total of 396 distinct toehold switches were thus designed for the PANTR1 lncRNA, some of them being common to several versions of PANTR1:
- 368 for the PANTR1 v1 (GenBank Acc. N° NR_037883)
- 38 for the PANTR1 v2 (GenBank Acc. N° NR_131233)
- 35 for the PANTR1 v3 (GenBank Acc. N° NR_131234)
- 44 for the PANTR1 v4 (GenBank Acc. N° NR_131235)
The CUHK tool has an embedded RNAfold algorithm that predicts the minimum free energy (MFE) of trigger, switch, switch-domains, and trigger-switch complex, and a ranking function that attributes an efficacy score to each toehold switch based on a linear regression model trained on 181 switches and their ON/OFF rates.
For PANTR1, we observed that one quarter of the designed toehold switches had predicted efficacy scores greater than 1 (Figure 2), and that the majority of them were specific to PANTR1 v1, which is statistically expected as the length of its sequence is greater than those of PANTR1 v2, v3 and v4.

Figure 2. Efficacy scores (ON/OFF rates) calculated by the CUHK model of the all switches
designed for PANTR1.
Among all the designed switches, we selected the top 3 best ranked and their cognate triggers to characterize them in vivo in E. coli, for a total of 8 switches and 10 triggers (Table 1) by taking into account the overlapping sequences between the four versions of PANTR1.
These 8 switches, designated hereafter by numbers, have slightly different secondary structures (Figure 3) and thermodynamic characteristics (Table 2). The minimum free energy (MFE) of the trigger-switch heterodimer is at least 25.1 and up to 43.2 kcal/mol lower than the total MFE of trigger and switch monomers. This value termed MFE difference in the CUHK model output represents the Gibbs free energy change needed for the RNA:RNA interaction to proceed. These 8 switches also have a high difference between the MFE of trigger-switch dimer and either of the trigger-trigger and the switch-switch dimers suggesting that the activated switch conformation is favored thermodynamically. Following these calculations, we expect that the switch n°8 will have beneficial result slightly higher than switch n°1 and that the switch n°7 will have lesser performance with any of the 3 cognate triggers. Comparing efficacy scores between genes, it is expected that the toehold switches designed to identify PANTR1 v1 lncRNA will have a much higher performance compared to those designed for the other three versions.
Table 1. Top 3 best ranked toehold switches designed by the CUHK pipeline for the four PANTR1
variants. This set of 8 toehold switches was later synthesized and evaluated in vivo in E. coli.
| Label | Targeted gene and rank number | |||
|---|---|---|---|---|
| PANTR1 v1 | PANTR1 v2 | PANTR1 v3 | PANTR1 v4 | |
| n°1 | Rank 1 | - | - | - |
| n°2 | Rank 2 | - | - | - |
| n°3 | Rank 3 | - | - | - |
| n°4 | Rank 23 | Rank 1 | Rank 1 | Rank 2 |
| n°5 | Rank 52 | Rank 2 | Rank 2 | Rank 4 |
| n°6 | Rank 58 | Rank 3 | Rank 4 | Rank 5 |
| n°7 | Rank 54 (trigger n°7c) | - | Rank 3 (trigger n°7a) | Rank 3 (trigger n°7b) |
| n°8 | - | - | - | Rank 1 |

Figure 3. Secondary structures of the top toehold switches represented in dot-bracket notation
coloured to match the different segments in Figure 1.
Table 2. Minimum free energy (MFE) of trigger, switch, switch-domains and trigger-switch
complex and efficacy scores estimated by the CUHK pipeline for the 8 PANTR1 toehold switches selected for in
vivo
characterization. Values are expressed in kcal/mol.
| Label | Minimum Free Energy (MFE) | Efficacy Score | ||||||
|---|---|---|---|---|---|---|---|---|
| RBS-Linker | Switch | Trigger | Trigger-Switch Dimer | Switch-Switch Dimer | Trigger-Trigger Dimer | MFE Difference | ||
| n°1 | -6.4 | -18.3 | -18.3 | -77.5 | -42.0 | -44.2 | 40.9 | 182.24 |
| n°2 | -5.8 | -28.0 | -19.3 | -84.3 | -60.0 | -50.2 | 37.0 | 140.84 |
| n°3 | -6.5 | -21.2 | -25.2 | -82.5 | -47.8 | -60.5 | 36.1 | 133.38 |
| n°4 | -6.4 | -26.2 | -27.9 | -85.9 | -59.1 | -64.5 | 31.8 | 74.68 |
| n°5 | -7.0 | -32.4 | -22.3 | -90.6 | -69.9 | -51.7 | 35.9 | 30.67 |
| n°6 | -4.9 | -36.1 | -21.6 | -92.0 | -77.3 | -48.4 | 34.3 | 25.61 |
| n°7a | -4.8 | -23.6 | -23.2 | -71.9 | -51.2 | -52.6 | 25.1 | 28.65 |
| n°7b | -4.8 | -23.6 | -22.8 | -72.8 | -51.2 | -50.2 | 26.4 | 37.62 |
| n°7c | -4.8 | -23.6 | -21.4 | -70.1 | -51.2 | -51.9 | 25.1 | 28.65 |
| n°8 | -6.6 | -28.5 | -28.3 | -100.0 | -63.5 | -64.7 | 43.2 | 79.61 |
Build
The 8 selected toehold switches sensors were assembled by Golden Gate technique in the low copy plasmid pSB3T5. For this, we designed and synthesized fragments containing the T7 promoter followed by the different toehold switches, the RBS Loop and the Linker sequences, with flanking type IIS restriction sites (BsaI) and inserted them into BBa_K3453101, which contains a Golden Gate adapter with BsaI sites upstream of the sfGFP-LVAtag.The 10 selected trigger sequences were placed under the control of the T7 promoter and followed by the strong SBa_000587 synthetic terminator. The resulting transcriptional units were then synthesized and assembled in the high copy plasmid pSB1C3.
The list and sequences of PANTR1 toehold switches and triggers is accessible through our Basic Parts, Composite Parts and Parts Collection sections.
Test
For toehold switch characterization, cells of E. coli BL21 Star™(DE3) (Thermo Fisher Scientific) were used. This strain was chosen, first of all because it contains inserted in its genome the T7 RNA polymerase gene, required for the transcription initiation of all PANTR1 toehold switches sensors and triggers. It is under the control of the lacUV5 promoter and requires IPTG to induce expression. And second because the it has the particularity to contain a truncated version of the RNAseE gene (rne131), which reduces the level of RNA degradation and thus increases RNA stability.E. coli cells containing both the toehold switch sensor and the trigger plasmids were grown overnight at 37 °C at 200 rpm in 96-deep-well plates with 1 mL of LB, in which was added 5 µg/mL tetracycline and 17.5 µg/mL chloramphenicol. The cells were then diluted by 40 times in the same media and after 4 hours incubation at 37°C at 200 rpm, they were further diluted by 20 times in media containing also 10 µM IPTG in an opaque wall 96-well polystryrene microplate (COSTAR 96, Corning). The plate was then incubated at 37°C at 200 rpm and the sfGFP fluorescence (λexcitation 488 nm and λemission 530 nm) and optical density at 600nm (OD600) were measured every 10 minutes for 24 hours, in a CLARIOstar (BMGLabtech) plate reader.
Fluorescence values were normalized by OD600. Furthermore, the arbitrary units have been converted into Molecules of Equivalent FLuorescein (MEFL) / particle (more details about the calibration curves are available on the Measurement page on this wiki).
Learn
Controls showed expected results
The results we obtained with the pairs of control (Figure 4) showed that the sfGFP expression was detected in the positive controls (custom made RBS and standard toehold switch RBS stem-loop) while there was no sfGFP expression detected in the negative controls (empty backbone and no promoter + no RBS), as expected.Three toehold switches significantly expressed sfGFP only in presence of PANTR1
Fluorescence results represented in Figure 4 reveals that among all the switches tested, 3 of them (n°5, 6 and 8) showed no fluorescence (or a negligible one) when the trigger was missing and a Fluorescence/OD600nm comparable to the positive controls when the trigger is present. This difference (i.e. the ON/OFF ratio) reaches a value of 102 for the toehold switch n°6 which is thus our best RNA sensor, followed by switches n°8 and n°5 with fold changes of 40 and 15 respectively (Figure 5).Three other switches (n°1, 3, 4) were able to express sfGFP to a level comparable to that of the positive controls in the presence of their cognate triggers, but, in the absence of the trigger, leakiness was observed. Their ON/OFF ratios (1.1, 1.5 and 2.4) are poor and were not further considered. Switches n°2 and 7 are not functional. No fluorescence is observed in any condition. The sfGFP expression is tightly controlled and the presence of the trigger does not allow the hairpin to unfold.

Figure 4. In vivo characterization of sfGFP expression by E. coli BL21
Star™(DE3)
cells
carrying the PANTR1 toehold switches and triggers. The negative controls have been performed with an empty pSB3T5
and BBa_K3453103 (no promoter, no RBS), and
pSB1C3 (no
trigger, “-” symbol) for every Switch. The positive controls have been performed with BBa_K3453104 (custom made RBS) and BBa_K3453105 (standard toehold switch RBS stem-loop). The data
and error bars are the mean and standard deviation of at least three measurements on independent biological
replicates.

Figure 5. MEFL/Particle fold changes of the PANTR1 toehold switches in the presence of their
cognate triggers compared to the MEFL/Particle value in the absence of the trigger.
Crosstalk
Additional crosstalk experiments were carried out to evaluate our 3 functional toehold switches behavior in the presence of other triggers.We thus tested switches n°5, 6 and 8 in the presence of the other 9 non-cognate PANTR1 triggers. Results presented in Figure 6 and 7 revealed that Switch n°8 showed significant fluorescence only with its cognate trigger n°8, while Switch n°5 and n°7 crosstalk:
- Switch n°5 showed a significant fluorescence with cognate trigger n°5 and non-cognate trigger n°6
- Switch n°6 showed fluorescence with trigger n°6 and a low fluorescence with non-cognate trigger n°5.

Figure 6. In vivo characterization of sfGFP expression by E. coli BL21
Star™(DE3)
cells
carrying the PANTR1 toehold switches n°5, 6 or 8 and all other triggers. The negative controls have been performed
with an empty pSB3T5, BBa_K3453103 (no
promoter, no RBS) and
pSB1C3 (no trigger), and the positive controls with BBa_K3453104 and BBa_K3453105. The data and error bars are the mean and standard
deviation of at least three measurements on independent biological replicates.

Figure 7. MEFL/Particle fold changes of the PANTR1 toehold switches n°5, 6 or 8 in the
presence of each of all the other triggers compared to the MEFL/Particle value in the absence of the trigger.
Efficacy scores of the CUHK model were different from in vivo toehold switches performance
The efficacy scores for PANTR1 toehold switches estimated by the CUHK model showed a different rank of efficacy than the one observed in vivo: Switch n°6 showed the best results in vivo, while Switch n°1 showed the best efficacy score in silico (Table 3). The efficacy score of the CUHK model alone was thus a poor predictor of in vivo efficacy of PANTR1 switches, in our case, as already described in the literature [8].Table 3. Efficacy scores of PANTR1 toehold switches designed by the CUHK tool (Figure 2) vs
their ranking according to fold change ratio measured in vivo (Figure 5).
| Label | Efficacy score in silico | Efficacy rank in vivo |
|---|---|---|
| PANTR1 Switch n°1 + Trigger 1 | 182.24 | 6 |
| PANTR1 Switch n°2 + Trigger 2 | 140.84 | 7 |
| PANTR1 Switch n°3 + Trigger 3 | 133.38 | 5 |
| PANTR1 Switch n°4 + Trigger 4 | 74.68 | 4 |
| PANTR1 Switch n°5 + Trigger 5 | 30.67 | 3 |
| PANTR1 Switch n°6 + Trigger 6 | 25.61 | 1 |
| PANTR1 Switch n°7 + Trigger 7a | 28.65 | 8 |
| PANTR1 Switch n°7 + Trigger 7b | 37.62 | 10 |
| PANTR1 Switch n°7 + Trigger 7c | 28.65 | 9 |
| PANTR1 Switch n°8 + Trigger 8 | 79.61 | 2 |
Conclusion
We have successfully designed, built, tested and analyzed 8 PANTR1 lncRNA specific toehold switches in E. coli. One of them (switch n°6) was strongly expressing the downstream sfGFP gene reporter in the presence of its cognate PANTR1 trigger and strongly repressing its translation in the absence of the trigger. It’s ON/OFF ratio of 101 highlights its performance and places it among the best switches described in the literature.Two others (switches n°5 and 8) were expressing sfGFP too, with a lower but very decent ON/OFF ratio.
These results can let us conclude the efficiency of 3 PANTR1 specific toehold switches in E. coli to detect the trigger and express a gene of interest in response, and thus the ability of our Electricia coli to act as a biosensor of PANTR1 lncRNA.
References
[1] Green AA, Silver PA, Collins JJ, Yin P. Toehold switches: de-novo-designed regulators of gene expression. Cell (2014) 159: 925–939.[2] Pardee K, Green AA, Takahashi MK, Braff D, Lambert G, Lee JW, Ferrante T, Ma D, Donghia N, Fan M, Daringer NM, Bosch I, Dudley DM, O’Connor DH, Gehrke L, Collins JJ. Rapid, low-cost detection of Zika virus using programmable biomolecular components. Cell (2016) 165: 1255–1266.
[3] Ma D, Shen L, Wu K, Diehnelt CW, Green AA. Low-cost detection of norovirus using paper-based cell-free systems and synbody-based viral enrichment. Synthetic Biology (Oxford, England) (2018) 3: ysy018.
[4] Suvanasuthi R, Chimnaronk S, Promptmas C. 3D printed hydrophobic barriers in a paper-based biosensor for point-of-care detection of dengue virus serotypes. Talanta (2022) 237: 122962.
[5] Park S, Lee JW. Detection of coronaviruses using RNA toehold switch sensors. International Journal of Molecular Sciences (2021) 22: 1772.
[6] Köksaldı İÇ, Köse S, Ahan RE, Hacıosmanoğlu N, Şahin Kehribar E, Güngen MA, Baştuğ A, Dinç B, Bodur H, Özkul A, Şeker UÖŞ. SARS-CoV-2 detection with de novo designed synthetic riboregulators. Analytical Chemistry (2021) 93: 9719–9727.
[7] Hunt JP, Zhao EL, Free TJ, Soltani M, Warr CA, Benedict AB, Takahashi MK, Griffitts JS, Pitt WG, Bundy BC. Towards detection of SARS-CoV-2 RNA in human saliva: A paper-based cell-free toehold switch biosensor with a visual bioluminescent output. New Biotechnology (2022) 66: 53–60.
[8] Soudier P, Rodriguez Pinzon D, Reif-Trauttmansdorff T, Hijazi H, Cherrière M, Goncalves Pereira C, Blaise D, Pispisa M, Saint-Julien A, Hamlet W, Nguevo M, Gomes E, Belkhelfa S, Niarakis A, Kushwaha M, Grigoras I. Toehold switch based biosensors for sensing the highly trafficked rosewood Dalbergia maritima. Synthetic and Systems Biotechnology (2022) 7: 791–801.
[9] Hong F, Ma D, Wu K, Mina LA, Luiten RC, Liu Y, Yan H, Green AA. Precise and programmable detection of mutations using ultraspecific riboregulators. Cell (2020) 180: 1018-1032.e16.
[10] To AC-Y, Chu DH-T, Wang AR, Li FC-Y, Chiu AW-O, Gao DY, Choi CHJ, Kong S-K, Chan T-F, Chan K-M, Yip KY. A comprehensive web tool for toehold switch design. Bioinformatics (Oxford, England) (2018) 34: 2862–2864.
[11] Huang A, Nguyen PQ, Stark JC, Takahashi MK, Donghia N, Ferrante T, Dy AJ, Hsu KJ, Dubner RS, Pardee K, Jewett MC, Collins JJ. BioBits™ Explorer: A modular synthetic biology education kit. Science Advances (2018) 4: eaat5105.